Light-harvesting complexes and Transmembrane protein studies
Photosynthetic conversion of the energy of sunlight into its chemical form suitable for cellular processes involves a variety of physicochemical mechanisms. At low light intensities, the quantum efficiency of photosynthesis is near unity, meaning that each of the absorbed photons reaches the RC and drives the electron transfer reactions. At high light intensities, however, the RC by regulation mechanisms that lead to quenching of excess excitation energy in light-harvesting proteins to mitigate oxidative damage. This process is known as non-photochemical quenching (NPQ), and it manifests itself by a strongly reduced chlorophyll fluorescence lifetime and by clustering and/or increased energetic interconnectivity of the light-harvesting antenna [J. Chmeliov, et al. J. Phys. Chem. Lett, 2019]. It is a common agreement that excitation quenching takes place in a specific pigment-protein complex of the light-harvesting antenna.
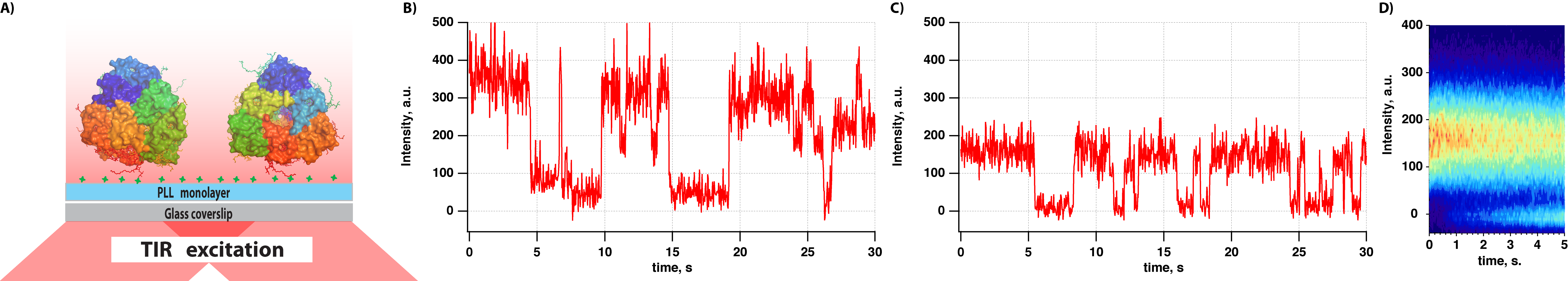
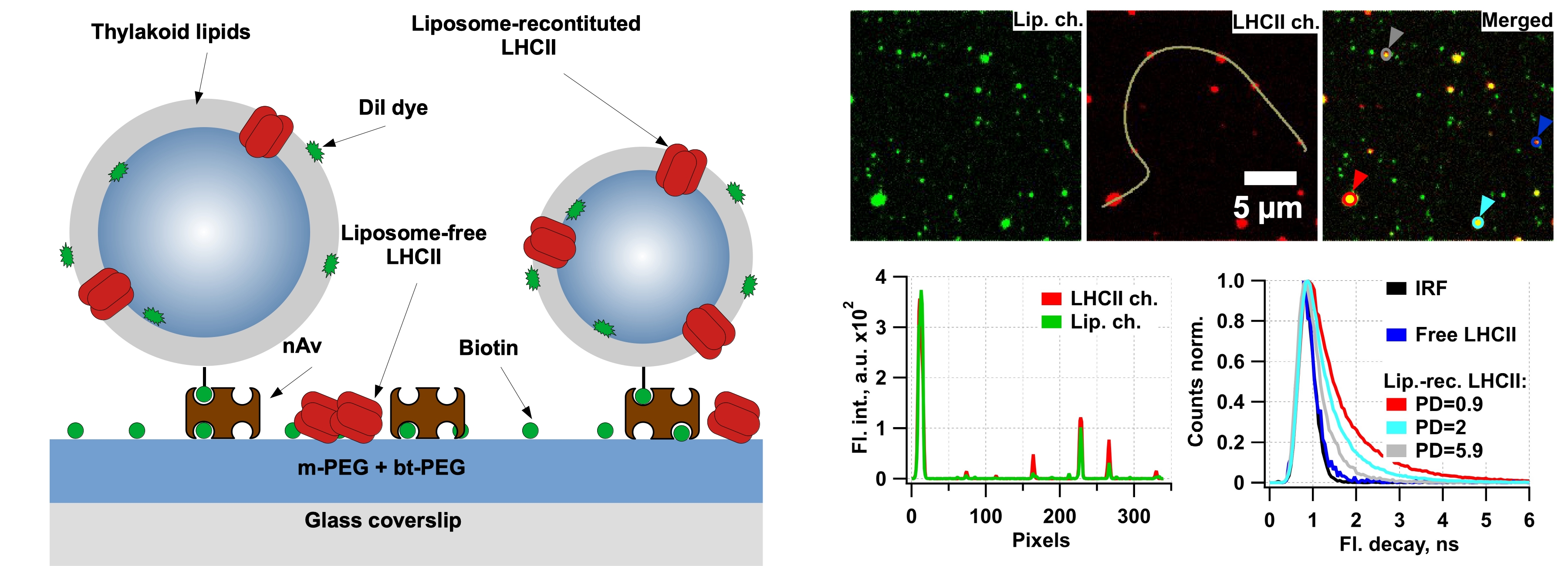
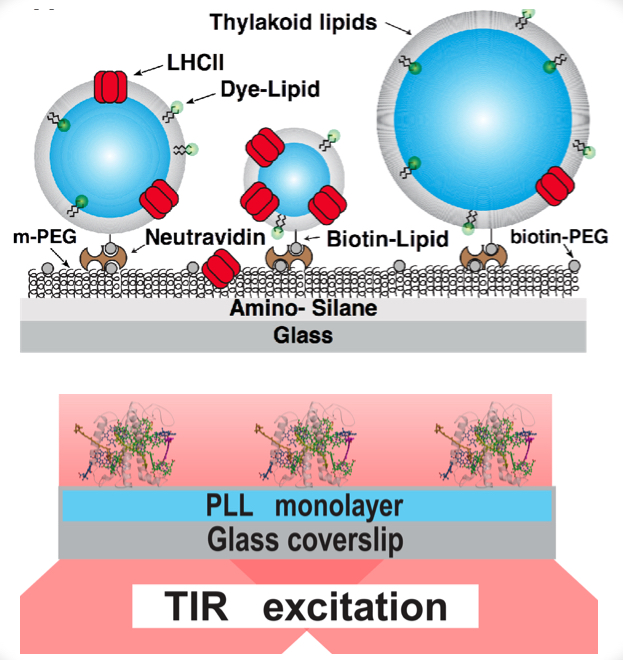
We perform studies of surface-tethered antenna complexes in detergent micelles, lipid vesicles or native chloroplasts. To sudy conformational dynamics (fluorescence intensity variation over time) we utilize single-molecule TIRF microscopy [M. Tutkus, et al. BBA - Bioenergetics, 2019], [M. Tutkus, et al. J. Phys. Chem. Lett., 2017]. To study quenching of fluorescence we also add the time-resolved equipment to our measurment set-up and then perform measuremtns in confocal mode [M. Tutkus, et al. Langmuir, 2018], [M. Tutkus, et al. J. of Photochemistry and Photobiology B: Biology, 2021]. This equipment is available at our department (Molecular Compounds Physics Department at the Center for Physical Sciences and technology) and it is based on a home-built fluorescence microscope set-up. This set-up has several constant-wave lasers, a pulsed red laser, dual-view and EMCCD camera, two avalanche photo-detectors and the time-correlated single photon counting card. This microscope also has an automated focus drift correction function, which enables long-lasting stable signal registration. Surface tethered antenna complexes in detergent micelles and their fluorescence intensity measuments at the single-molecule level. Antenna complexes in surface tethered small unilamelar vesicles and their fluorescence life-time measuments at the single-liposome level. We reconstitute transmembrane proteins (the same applies for membrane-related proteins) in lipid vesicles (small unilamellar vesicles - SUV) supplemented with hydrophobic membrane stain and biotinylated lipids. Next, we immobilize the proteo-liposomes sample on the PEGylated glass coverslips via streptavidin or similar protein. Membrane dye allow us to perform co-localization between antenna complexes (635 nm excitation) and membrane (532 nm excitation) fluorescence channels. This co-localization strategy helps to discriminate between SUV-reconstituted and SUV-free antenna complexes bound to surface. Also, membrane dye fluorescence intensity allows intensity-based characterization of vesicle size, which is an interesting parameter for antenna complex quenching characterization (e.g. whether LHCII in smaller lipid area are more quenched).Antenna complexes in surface tethered small unilamelar vesicles (top) and detergent micelles (botttom) To compare role of antenna complexes in the non-photochemical quenching process both under native and artificial lipid environment we perform ensemble level time-resolved fluorescence temperature-dependence measurements of antenna complexes in native thylakoids. We use streak-camera and kryostat for these measurements. The streak-camera reveal wavelength-dependency of LHCII fluorescence lifetime. At the same time, sample is mounted on the kryostat, which enables cooling of the sample to 15 K temperature.
These experiments help us to answer the following questions: a) discriminate vesicle-free/vesicle-reconstituted antenna complexes and estimate antenna complexes reconstitution efficiency. b) find, if existing, fluorescence kinetics differences between vesicle-reconstituted (aggregates in vesicle membrane) and vesicle-free (aggregates bound to surface) antenna complexes. c) whether different antenna complexes vesicles (different number of antenna complexes per similar area vesicle or similar antenna complexes number per different vesicle area) has influence on the antenna complexes fluorescence kinetics. d) whether vesicle size alone (despite number of antenna complexes per vesicle) has an influence on antenna complexes fluorescence kinetics. e) find differences, if existing, between antenna complexes quenching under native and artificial lipid environment.